The metal-binding domain contains an unusual metal receptor site in which the bound metal is inaccessible to the solvent. The metal ion is bound with a linear coordination to two conserved cysteine residues. This dithiolate coordination confers the metal ion selectivity for +1 ions
[11]. The sensitivity of CueR for copper(I) ions is extremely high (in the zeptomolar range), which corresponds to less than one atom per cell. Apparently it is the task of CueR and CopA to clear the toxic copper ions rapidly from the cytoplasm into the periplasmic space
[11]. It has been demonstrated that up to two Hg
II ions bind with high affinity to WT CueR, one at the functional metal binding site (C112 and C120) and the other at the C-terminal CCHH motif. This motif provides a selection of ligands that could be participating in coordination of both soft and intermediate metal ions
[12]
The CueR residues S4, K15, F19, Y20, N34, and Y36 appear to be essential to activate transcription
[1]
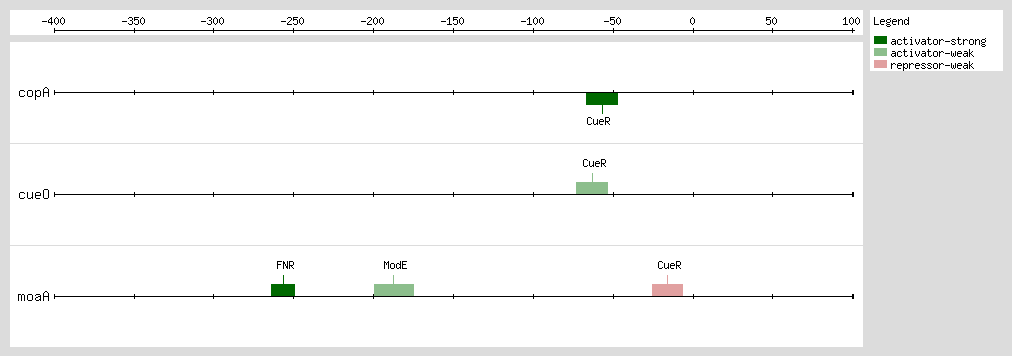
CueR activates transcription by overlapping the -35 box of promoters and represses it by binding downstream of the transcription start site
[6]. CueR binds to a 19-bp sequence containing two 7-bp inverted repeats separated by a 7-bp spacer
[6]. Binding occurs
in vitro in both the presence and the absence of copper ions. Holo-CueR binds more strongly to DNA than apo-CueR and with a higher conformational flexibility
[13]. Both holo- and apo-CueR bend DNA
[13]. The RNA polymerase is capable of binding to the
copA promoter independently of the holo or apo conformation of CueR, generating two types of interactions between RNA polymerase and the
copA promoter: one is favored by the holo-CueR conformation to activate transcription, and the other one is favored by apo-CueR to repress transcription
[14] Holo-CueR is able to change the structure of DNA around the promoter, allowing transcription initiation
[1] The unbinding process between protein-DNA interactions was studied using CueR as the model
[15] The formation of permanganate-sensitive transcription complexes requires copper ions
[6].
Based on electron paramagnetic resonance (EPR) spectroscopy, various active state conformations of the transcriptional regulator CueR were detected
[16] The structure of CueR in the active state was resolved, and this information was highly useful in identifying a specific site for inhibition and the critical role of α-helix 4 (α-4) in the transcription process
[16] The potential uses of double-histidine (dHis)-based Cu(II) spin labeling for identifying various conformational states of CueR during the transcription process were shown
[16]
CueR, a transcriptional dual metalloregulator, is subject to ATP-dependent proteolysis in the presence of Lon and ClpXP and ClpAP machineries
[17]. Its degradation is not altered by mutations affecting copper binding, dimerization, or DNA binding, but it requires an accessible C terminus
[17].
Reviews:
[18, 19, 20, 21, 22]