Binding of Co(II) and Ni(II) to RcnR affects its flexibility at the N terminus, and this effect modulates its DNA-binding affinity
[7]
The
rcnR gene is encoded divergently to the
rcnA gene, which encodes an efflux protein that is involved in nickel and cobalt homeostasis
[6]; it removes an excess of ions
[1]. It has been proved by Koch and colleagues
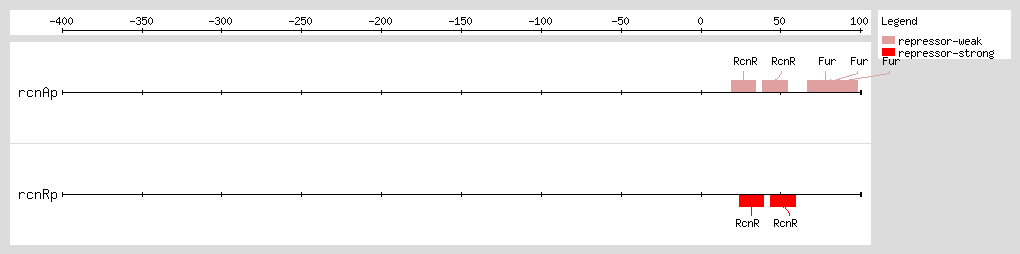
[2] that the
rcnR gene is the target of Fur regulation in regard to iron levels (when iron increases,
rcnR expression increases about 2.3-fold
[2]).
An RcnR structure study showed that it has a charged N-terminal region (amino acids 1-34), followed by a highly conserved region of primarily nonpolar amino acids (35-64), that is, a region probably involved in binding metals. The N terminus determines metal-specific responses via recognition of cognate and noncognate metals
[8]. Replacement of the Arg-14 and Lys-17 residues on RcnR affects its DNA-binding affinity
[7] Its secondary structure prediction shows three helices without any β-sheet
[1].
Cognate metals bound to EcRcnR in six coordinate sites composed of (N/O)
5S ligand donor atoms, while noncognate metals showed lower coordination numbers
[9].
RcnR is not similar to any other known transcriptional regulator; instead, it is homologous to other proteins whose genes are encoded next to genes encoding putative metal efflux proteins, metalloenzymes, or copper chaperones [and just a few (13%) of the homologs to cardiolipin synthases or membrane proteins]
[1]. Such conserved localization with genes of metal-related functions suggests that these homologs are likely to play a regulatory role in metal homeostasis
[1].
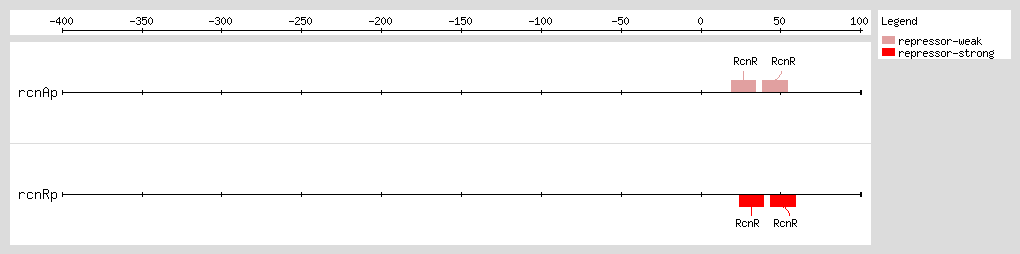
RcnR acts as a transcriptional repressor for its divergent
rcnA gene
[1] and a weak repressor for its own expression
[4]. In response to increasing concentrations of nickel or cobalt, RcnR releases the repression of
rcnA and
rcnR to allow its expression
[1, 4]. As
rcnR expression is induced in response to iron even in the absence of Fur
[2], it is possible that RcnR responds to iron ions directly.
RcnR binds to two sites in the
rcnA-rcnR intergenic region with the sequence TACT-G(6)-N-AGTA
[1, 4]. The TACT inverted repeats are critical for specific binding. The G-tracts provide a structural component to facilitate binding of RcnR to DNA, because they endow A-form DNA characteristics to the RcnR operators. In addition, RcnR interacts nonspecifically with the approximately 50 bp flanking the core binding site, resulting in DNA wrapping and the introduction of a single negative supercoil into plasmid DNA
[4]. RcnR forms a stable tetramer and binds as a tetramer to one operator. The tetrameric state is not perturbed by Ni(II) or Co(II) binding
[9].
Based on a computational model of apo-
EcRcnR in the tetrameric state, using experimental data it was determined that the model shows a large number of putative metal-binding residues localized at the interface between two monomers in the vicinity of Cys35 from one chain; the N-terminal amine and the imidazole rings of His3, His64, His67, and His60, known to be involved in metal binding, are all found in this region. In addition, Glu34 and Glu63 are also located in this potential metal-loading region, and their carboxylate groups could act as ligands for Co(II) and Ni(II)
[10].
Crystallization of RcnR-DNA complex with DNA that encompasses the DNA-binding site and X-ray diffraction data to 2.9 angstroms resolution were determined
[11].
X-ray
absorption
spectroscopy (XAS) analysis confirmed that both the Glu34 and Glu63 are ligands in the Co(II) complex, while for Ni(II), only Glu63 unambiguously acts as a ligand
[10]. Cys35 appears to be a ligand for Co(II), but not for Ni(II), while His60 is not involved in Ni(II) binding and is only slightly involved in Co(II) binding
[12].
As nickel levels depend on the Nik uptake system (NikR regulator, NikABCDE transporter) and nickel is highly used during anaerobic growth by the Ni-Fe hydrogenase isozyme, the additional participation of RcnR and RcnA in nickel homeostasis changes the previous understanding of the cell's nickel processing. Iwig and colleagues
[1] showed that RcnR is not directly related to other proteins that use nickel, although they showed that RcnA, as the exporter of excess ions, has direct participation. RcnA likely competes with NikR for nickel ions at low concentrations, and in such a way RcnA downregulates
nikABCDE expression and nickel uptake (for this,
rcnA must be expressed at a basal level prior to induction, as observed with higher nickel levels)
[1]. Also, RcnA is important in the modulation of the hydrogenase assembly pathway through the NikR regulation that limits nickel delivery to the process, repressing
nikABCDE transporter expression when intracellular nickel levels are high
[1].
Review:
[13]