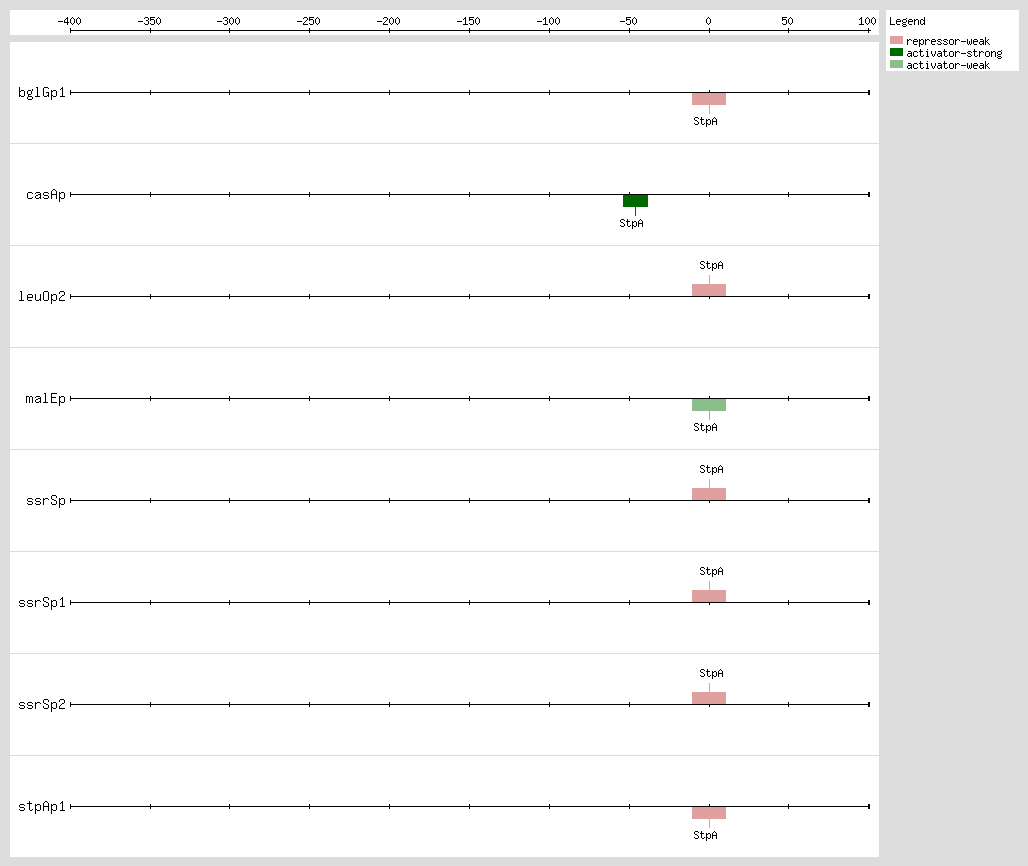
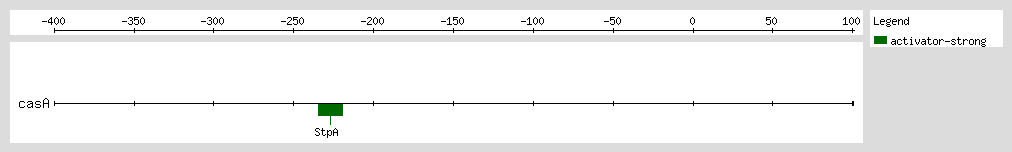
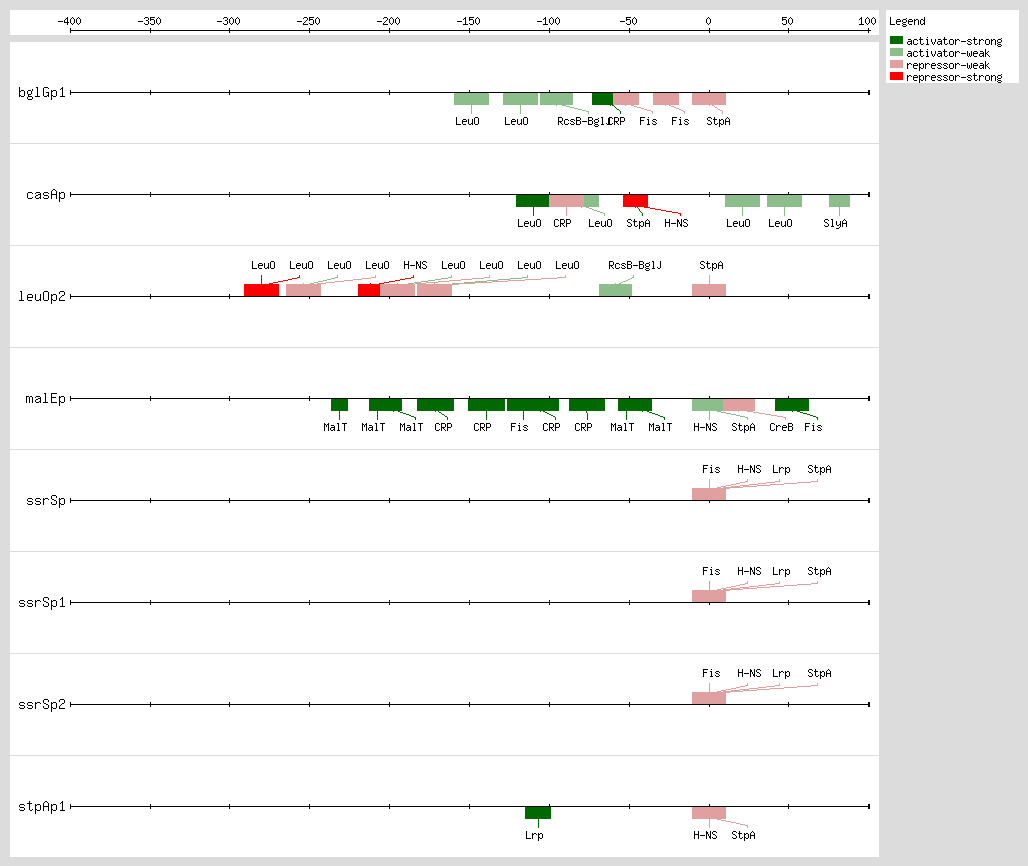
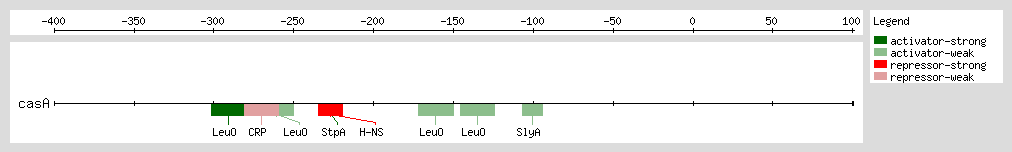
Gueune et al. suggested that StpA may regulate the expression of a tributyltin-inducible operon
[1]. Currently, no inducer for this regulator has been reported in the literature. New genes may be identified by high-throughput analysis
[18].
It is a DNA-binding protein with similarity to H-NS
[19, 20], and these two proteins can have similar functions
[10, 21]. It has an approximately fivefold-greater affinity for DNA than H-NS and has a preference for curved DNA
[22]. This regulator binds curved DNA, in the promoter region, with AT-rich motifs, and StpA binds to sites that have poorly related sequences and it does not have a consensus sequence for the DNA binding.
Expression of
stpA from a plasmid can complement an
hns mutant phenotype and StpA is able to repress and activate a subset of H-NS-regulated genes, but their specific mechanisms remain to be determined
[18, 19, 22, 23, 23, 24]. A dominant negative form of StpA can disrupt H-NS activity and vice versa, and H-NS can interact with StpA at two distinct domains to form heterodimers in vitro; also there exists evidence that some of these proteins can form homodimers
[25, 26, 26, 27]. For this reason, in the absence of H-NS the StpA protein is rapidly degraded in a Lon protease-dependent manner
[25, 28]; protection from proteolytic degradation appears to be mediated by direct interaction between StpA and H-NS
[25]. On the other hand StpA may form heteromeric complexes with Hha and YdgT
[29]. StpA and Hha stimulate pausing by RNA polymerase at 20
oC by promoting DNA-DNA bridging of H-NS filaments, through H-NS-mediated gene regulation
[30].
StpA consists of two structured domains which are separated by a protease-sensitive linker
[31]. The N-terminal domain is involved in the protein-protein interaction, while the purified C-terminal domain alone promotes annealing of RNA oligonucleotides and
trans-splicing in vitro
[26, 27, 31, 32]. Repression of the
bglG operon by StpA occurs as a molecular adapter
[15, 16] and only in the presence of the N-terminal H-NS protein domain, which comprises the core for dimerization
[11].
StpA has RNA chaperone activity in vitro and in vivo, and the RNA chaperone activity of StpA is influenced by the structural stability of the RNA target molecule
[14, 24, 32, 33, 34, 35, 36]. StpA may destabilize the small regulatory RNA MicF
[37, 38].